植物オルガネラ型DNAポリメラーゼ (POP)
今回の研究では,
植物オルガネラ局在型DNAポリメラーゼ(POP)をはじめて精製し,その性質を調べま
した.
FEBS Journalに論文が受理されました.
ミトコンドリアや色素体には,独自のDNAが存在し,これらは専用のDNAポリメラーゼによって複製されます.
動物(ヒトやマウスなど)や菌類(イーストなど)の場合,40年以上前からガンマというポリメラーゼが知られています.初期の研究で,真核生物からは3種
類のDNAポリメラーゼが精製され,アルファ,ベータ,ガンマと名付けられました.アルファは,その後発見されたデルタなどとともに核ゲノムの複製に働き
ますが,ベータは核の修復系で働くことがわかりました.ガンマはミトコンドリアの複製酵素であることが昔からわかっていて,ミトコンドリアの複製酵素は一
般的にガンマであると
信じられてきました.2000年になって植物の一種であるシロイヌナズナのゲノム配列が公表されると,ガンマがないことが分りました.また,それ以前から
の研究で,植物の色素体とミトコンドリアにあるDNAポリメラーゼは,大腸菌のDNAポリメラーゼI(PolI)と似ていることが指摘されていました.

図1:さまざまなDNAポリメラーゼの構造の比較
オレンジがDNAポリメラーゼドメイン,ピンクが5'-3'エキソヌクレアーゼドメイン,水色が3'-5'エキソヌクレアーゼドメイン。
そこで,改めて,植物のゲノムを調べてみると,PolI類似の酵素をコードすると思われる遺伝子がいくつかみつかりました.このうち2個については,ポリ
メラーゼドメインと3'->5'エキソヌクレアーゼドメインをもつものの,本来のPolIに存在する5'->3'エキソヌクレアーゼドメイン
は存在しないことが分りました.どちらも葉緑体輸送配列と思われる延長配列がN末端についていることが分り,その後2004年になって,GFP融合タンパ
ク質を用いた輸送実験により,どちらも葉緑体とミトコンドリアの両方に輸送されることが示されました.他方,イネのEST(expressed
sequence tag:
cDNAと同じ)からも同様の配列がみつかり,これをもとに完全な配列が得られOsPolI-likeと名付けられました.これを大腸菌で発現した酵素
は,確かにポリメラーゼ活性と3'->5'エキソヌクレアーゼ活性をもつことが分りました.さらに,抗体を用いたブロッティングにより,イネの芽生
えから得られた葉緑体画分にこの酵素が含まれることも分りました.
2004年に発表された単細胞性紅藻Cyanidioschyzon merolae(シアニジオシゾン,略してシゾンとよぶこともある)
のゲノムからも,PolIに類似した配列がみつかりました.それらをPolA,
PolBと呼ぶことにしました.PolAは,PolIと同様,3つのドメインを含んでいました.また,系統解析から,細菌の酵素と同じ中間であり,恐らく
水平移動によりシアニジオシゾンのゲノムに移ってきたものと考えられます.一方,PolBの方は,植物がもつPolI-likeと似ていて,5'-
>3'エキソヌクレアーゼドメインをもっていませんでした.さらに,葉緑体輸送配列と考えられるN末端の延長配列もありました.
詳しい系統解析の結果を図2に示します。
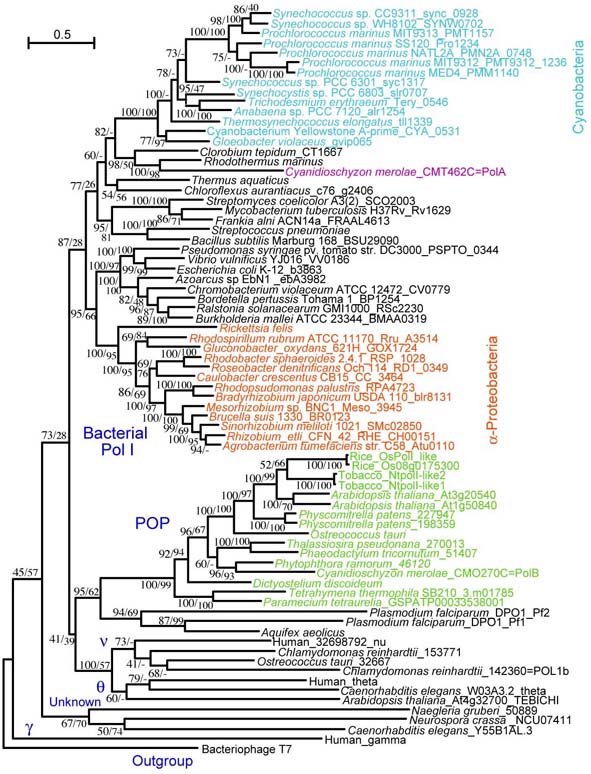
図2:細菌のDNAポリメラーゼ(Pol I)と植物や藻類のオルガネラ局在型DNAポリメラーゼの系統樹
TreeFinderを用いた最尤法による系統樹で,分岐の数字は左が最尤法,右が,近隣結合法(MEGA 4)による信頼度を表すパーセント。
PolBや上記の植物の酵素は1つのグループをつくっていること,さらに,このグループは細菌のPolIのクレードの外にあり,姉妹群と考えられることが
判明しました.はじめに述べたガンマはこれ等とは大きく隔たった酵素であることも明らかになりました.更に驚いたことに,動物・菌類(オピストコント)と
植物・藻類(バイコント)が分れる前にこれらから分れたアメーバ類や粘菌類にも,植物・藻類と同様のPolI-likeタイプの酵素があることが分りまし
た.こうした結果は,これらの酵素がPolIとは異なる捌のグループの酵素であり,真核生物の誕生の時からミトコンドリアの複製に使われていて,その後,
シアノバクテリアの共生によって藻類が誕生した時には葉緑体の複製にも使われるようになり,藻類から陸上植物が進化したのちにも保存されているということ
が考えられます.これに対し,動物や菌類の共通祖先が産まれたときに,ミトコンドリアの複製酵素が,外来の恐らくはウィルス由来のDNAポリメラーゼに
よって置き換えられて,現在知られるガンマになったと考えられます.後に述べる様々な証拠から,植物・藻類・アメーバにあるオルガネラ複製酵素に対し,
Plant organellar polymerase (POP)という名称を使うことを提案しました.
これまでに,POPの研究はさまざまにおこなわれてきていますが,実際に細胞からPOPを精製した研究はありませんでした.それは,植物細胞の場合,実際
に葉緑体やミトコンドリアの複製がおこなわれているのが分裂組織に限定されていて,複製酵素を高濃度に含む材料が得られなかったこと,植物は比較的低い温
度で育つ生物であるため,酵素の安定性もあまり高くなく,完全な形で精製することがむずかしいこと等の理由によると考えられます.この点,紅藻シアニジオ
シゾンは,46℃というかなり高い温度が至適であるうえ,単細胞生物なので,活発に分裂増殖をしている細胞を大量に得ることができます.そこで,研究室の
森山崇君は,シアニジオシゾンの細胞から,PolBを約600倍精製し,ほぼ純粋な酵素標品を得ることに成功しました.この酵素は,ポリメラーゼ活性と
3'->5'エキソヌクレアーゼ活性を示しました.合成されるDNAはかなり長く,5分間の反応で,1.3
kntにまでなりました.組み換えタンパク質として精製されたPolAは70
nt程度までしかDNA鎖を合成しなかったので,PolAは細菌のPolIと似ている一方,PolBは高いprocessivityをもつということが分
りました.Processivityというのは,一度鋳型に結合したポリメラーゼが,鋳型からはなれることなく,どのくらいのDNAを合成するかという尺
度で,ゲノムの複製に働く酵素ではprocessivityが高く,修復酵素では低いのがふつうです.これまでに組換え体として研究されたイネのPOPに
比べても,高いprocessivityが認められたことは,細胞から精製したnativeな酵素だからだと思われます.
PolBのN末端配列をGFPにつないだキメラ遺伝子を,タマネギの細胞に導入し,GFPの局在を調べたところ,色素体に局在することが分りました.シア
ニジオシゾンで同じ実験が出来ればよいのですが,まだシアニジオシゾンで形質転換をおこなうことはかなりむずかしいため,このようなやり方をしています.
それでも,シアニジオシゾンのいろいろな色素体タンパク質について,同様に調べたところ,期待どおりの結果が得られているので,紅藻と植物で,色素体への
輸送シグナルは共通していると考えられます.これは,ゲノム情報解析により,良く似た輸送装置が検出されていることとも対応します.
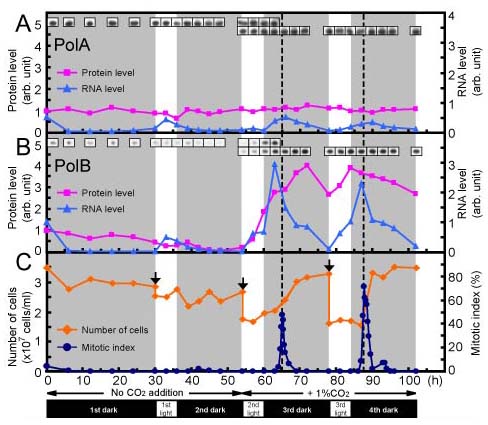
図11:細胞周期におけるPolA, PolBの発現
では,PolBがほんとうに複製に働いているのでしょうか.これに対する直接的な証明はかなり困難ですが,同調培養系を用いた実験により,細胞が分裂する
こととPolBの発現が関連していることがわかりました.この同調系はあたらしく開発したもので,空気をとおして培養したときには6時間の明期があっても
細胞は分裂しませんが,1%の二酸化炭素を含む空気をとおした場合には,明期のあとで細胞が同調して分裂します.このようにして,明暗サイクルは同じで
も,二酸化炭素の有無で,細胞が分裂したりしなかったりするという系を開発しました.この結果,分裂しないサイクルでは,PolBは発現していませんが,
分裂するサイクルでは,M期にPolBの遺伝子が発現することが分りました.これは次のサイクルで必要とされる酵素を補充するためだと思われます.
PolBがオルガネラの複製に働いていることを直接証明するには,遺伝子破壊株を利用することが必要になりますが,シアニジオシゾンの遺伝子破壊や
RNAiはまだ開発されておらず,今回の研究では利用することが出来ませんでした.そのため,間接的ではありますが,分裂との関係が得られたということ
で,PolBがオルガネラの複製に関わる可能性が示されたと考えています.
この他のデータも含め,今回の一連の研究により,シアニジオシゾンのPolBが植物・藻類に共通なオルガネラ複製酵素であることが分りました.
Last update: April 2, 2008.
Copyright © 2008
Sato Lab. All Rights Reserved.